
Mycorrhizal fungi: Invisible aid
By PD Dr Christel Baum, Faculty of Agricultural and Environmental Sciences, University of Rostock
Interest grows in alternative strategies for crop production: approaches that differ from the classic reliance on fertiliser and plant protection. Biostimulators can boost plant growth, although there’s need for much more research. Mycorrhization featuring symbiosis of fungi and plant roots is a well-known phenomenon in this respect. Mycorrhizal fungi are constant companions of most crops, making important contributions to plant nutrition, crop health and soil fertility. Particularly where P fertiliser application is reduced, the importance of mycorrhization in plant nutrition increases. Mycorrhizal fungi can help the utilisation by plants of already existing soil P reserves without inoculation being necessary.
Multiple benefits
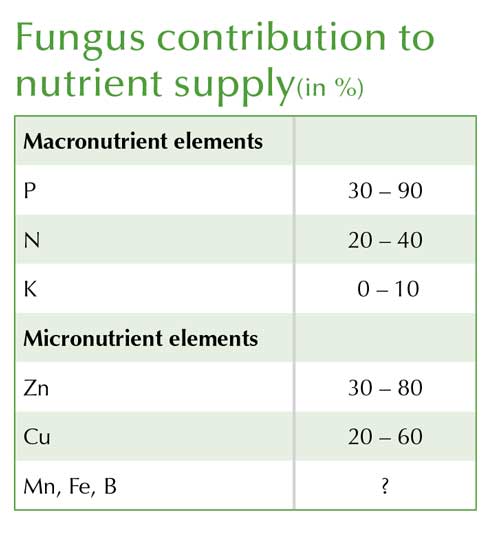
Worldwide, around 300 species of the relevant (Glomeromycota) fungi have been described. On any one location of crop growing land only between 15 and 25 species may be found in most cases, although these will be able to colonise all suitable host plants. Compared to many other symbiotic organisms, these fungi are less host-specific. One host plant can be colonised by several species of mycorrhizal fungi simultaneously. The beneficial effect of an individual fungus species for the host plant can vary substantially while the overall effect usually includes improved supply of phosphate, nitrogen, sulphur and often also of zinc and copper. The mycelium or hyphae network of mycorrhizal fungi can increase by a factor of five the area from which the host plant can access nutrients with moisture supply also improved, boosting drought tolerance of the host plant.
How does this symbiosis function?
Mycorrhizal fungus spores are very large compared with those of other fungi with some almost 1 mm in diameter. Root exudates from suitable host plants encourage spore germination and hyphae development. During mycorrhization, the fungus hyphae grow in the fine roots of the host plants, developing so-called »arbuscles« (strongly-branched fungus hyphae) in the root outer cells as sites for substance exchange between host plant and fungus partner. Mycorrhizal fungi also associate with so-called »mycorrhization support bacteria«. These extend the capabilities of the mycorrhizal fungi, encourage hyphae growth and mobilisation of nutrients.
Mycorrhizal fungi host plants
All sweet grasses (Poaceae) and most legumes (exceptions include lupins and ornithopus), as well as most other crop species, are hosts for mycorrhizal fungi. Not colonised by mycorrhizal fungi, for instance, are cruciferous plants such as oilseed rape and oil radish. Including non-host crops in rotations reduces mycorrhizal fungi colonisation intensity, although not sustainably so. Fungus spores remain viable over several years and can often find host plants amongst weed populations, although the major proportion of the typical seed-propagated weeds e.g. Chenopodium or goosefoot and knotweed species, show a limited to non-existent affinity to mycorrhization.
Plants not colonised by mycorrhizal fungi compensate for lack of mycorrhization aid in nutrient intake through a more extensive root system as well as more, and longer, root hairs. Additionally, numerous non-mycorrhizal plants exude into the soil allelopathic substances in order to deter settlement of mycorrhizal fungi and other plant species. This effect causes a temporary reduction in density of mycorrhizal fungi spore population after growing such crops, although no permanent damage in this respect.
Can breeding improve plant mycorrhization?
Although mycorrhization has been demonstrated in the majority of crops, its intensity can differ substantially, even between different varieties of the same crop, indicating a perspective for breeding crops for improved symbiosis: a target being looked into at the moment, although nothing has reached variety testing so far. As well as plant phenotype influencing mycorrhization affinity of host plants, crop development stage also has an influence. For this reason, there is no established recommended mycorrhization rate for fine roots, instead only an empirical crop-specific general value based on the proportion of fine root lengths colonised by mycorrhizal fungi (e.g. approx. 50% for maize).
A high mycorrhization rate for roots is seen as indicator of increased fungus efficacy in terms of encouraging host plant growth. Basically, mycorrhization can have a growth promoting, a growth neutral or even, in individual cases, a growth limiting effect on host plants. Both partners in this symbiosis, fungus and plant, can regulate the exchange of exudates. Where the behaviour of a partner is parasitic, the other can retaliate by reducing transfer of its own exudates. Hereby, mycorrhization can tend towards neutrality in promoting plant growth where a good supply of nutrients is already available in the soil, and often towards growth promotion where there’s scarcity of nutrients. Non-host plants, and even host plants, can have their growth inhibited where there’s excess colonisation density of mycorrhizal fungi. But because this is a very rare phenomenon documented in just a single study, there’s no need for controls or regulation in that the advantages of mycorrhization exceed the possible disadvantages by a long way.
Improved disease resistance
Along with the effect on nutrient supply, colonisation by mycorrhizal fungi can lead to physiological changes in the host plant and also to induced systemic resistance to disease and pests. The colonisation of the roots by mycorrhizal fungi starts off the same chain of signals as does infection of the plant by a pathogen. This is why resistances can emerge in parts of the host plant that are not colonised (systemic acquired resistances). These are activated by semiochemicals or messenger substances distributed over the entire plant, preparing it physiologically for pathogen attack. Important semiochemicals in this respect are jasmonic acid and salicylic acid. Volatile semiochemicals can also be spread by wind, in this way also activating neighbouring plants and increasing development of secondary metabolites for defence against diseases and pests.


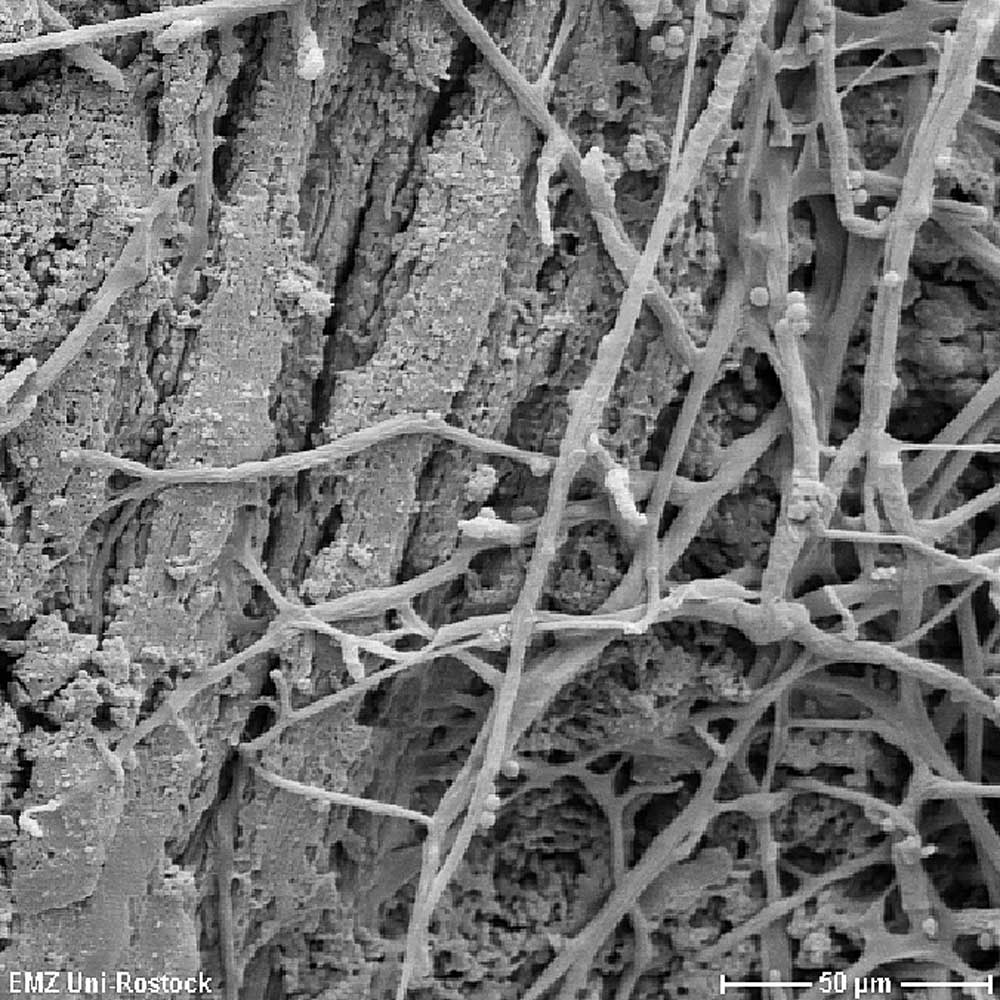
Effects on soil structure
Mycorrhizal fungi have an important soil ecological function in creating and stabilising the top soil crumb structure. Stabilisation is achieved mechanically through the hyphae network and chemically via fungus protein production. In this way, carbon sequestration in soil is encouraged, as well as aeration improved and moisture holding capability increased.
Mycorrhizal fungi can develop downwards for over 1 m into the subsoil, improving plant access to nutrient reserves there.
How can the farmer help?
In fact, many agricultural operations limit the beneficial actions of mycorrhizal fungi, especially work such as cultivating and fertiliser application. Working the soil (particularly inversion methods) weakens the hyphae network through mechanical damage, interrupting nutrient and water transport channels and drying-out hyphae. Fertilising operations, particularly application of P, reduces the reliance of host plants on mycorrhization, encouraging more self-sufficiency in both symbiosis partners rather than mutually advantageous association. Result: the intensity of mycorrhization reduces.
Conclusion
With reduced application of fertiliser, the benefits of mycorrhization increase in crop production. Realistic expectations from a vital mycorrhization in a growing crop are not in the first line yield increases but instead increased harvest security and quality. Effective measures encouraging mycorrhization include reduced soil cultivations as well as growing of perennial crops and catch cropping. Such strategies encourage spore production and therefore increase spatial colonisation potential. In the long term, plant breeding could play a role through producing new varieties with increased specific mycorrhization affinity for stronger benefits in crop production.
What can biostimulators offer?
Biostimulators based on live microorganisms, including mycorrhizal preparations, are influenced in their efficacy by many environmental factors. For this reason, a precise prognosis of their efficacy in the open field is barely possible. Basically, spatially specific chemical and physical soil characteristics (above all, crumb structure, pH and organic content) have a formative influence. However, weather during and after application, as well as the soil’s own microbiome, also have decisive influence on establishment, expansion and efficacy of biostimulators in the form of live mycorrhizal preparations.
Such preparations are effective in soil and substrates that do not have their own inoculation potential. In particular, this covers substrates in horticulture and damaged substrates, e.g. in landscapes being reconstituted following open cast mining, or in urban areas being prepared for recultivation. Numerous mycorrhizal preparations, often containing spores on expanded clay substrate, are available for supporting mycorrhization. On the other hand, in arable fields the soil’s own inoculation potential should be exploited. The spore population density of mycorrhizal fungi as indicator of the inoculation potential can be determined through wet sieving and microscope spore counting. On locations without damaged substrate, sufficient inoculation potential may be assumed without testing.
Growing catch crops encourages spatial inoculation potential of soil through increasing mycorrhizal fungi spore population density, especially where the catch crops have a high mycorrhization affinity. This applies to crops such as phacelia and green rye and also seed mixes with clover, strigose oats and vetches.
